Spletna revija za znanstvenike, strokovnjake
in nevroznanstvene navdušence
Naslovnica Članki Intervjuji Mnenja Zdravje Korenine eSinapsa Številke ![]()
Branje in možgani
članki
eSinapsa, 2011-1
Zvezdan Pirtošek
Eksoskeleti – inteligentne bionske naprave
Marko Munih
O aktualnih dilemah draženja globokih možganskih struktur pri obsesivno - kompulzivni motnji
Nadja Jarc
Sledite svojo srečo ... z iPhone
Urban Kordeš
eSinapsa, 2011-2
Renata Salecl
Gašper Tkačik
Astrociti – spregledane zvezde nevrobiologije
Marko Kreft, Robert Zorec
Sašo Dolenc
Meditacija - malo truda, veliko koristi
Luka Dimic
eSinapsa, 2011-3
Mara Bresjanac
Martina Starc
Rok Berlot
Varnost uporabe generičnih protiepileptičnih zdravil
Mojca Kržan, Matevž Kržan
Možgani, računalniki - nekaj vmes
Miha Pelko
eSinapsa, 2012-4
Ali so moški in ženski možgani različni?
Gregor Majdič
O kognitivnih motnjah pri bolnikih s Parkinsonovo boleznijo
Dejan Georgiev
Akutno možgansko kap lahko uspešno zdravimo
Nina Vujasinovič, Bojana Žvan
Vloga nevropsihološke diagnostike pri odkrivanju zgodnjih znakov alzheimerjeve bolezni
Simon Brezovar
eSinapsa, 2013-5
Novo odkritje na področju sporadičnih prionskih bolezni
Jana Jerše, Nadja Jarc
Učinek placeba brez lažnih zdravil in zavajanja
Mara Bresjanac
Subarahnoidna krvavitev zaradi tromboze venskih sinusov
Mateja Repar, Anita Resman Gašperčič
Srečanje dveh velikanov: možganov in imunskega sistema
Matej Markota
eSinapsa, 2013-6
Odstranjevanje možganskih tumorjev pri budnem bolniku
Andrej Vranič, Jasmina Markovič, Blaž Koritnik
Zmedena bolnica, ki nič ne vidi ali PRES
Manja Hribar, Vid Zgonc
Manja Hribar
Netravmatska lokalizirana konveksitetna subarahnoidna krvavitev
Mateja Repar, Fajko F. Bajrović
Sistemska skleroza in ishemična možganska kap - vzročna povezanost ali le koincidenca?
Mateja Repar, Janja Pretnar Oblak
Klemen Grabljevec
Z omejevanjem spodbujajoča terapija pri bolnikih po nezgodni možganski poškodbi
Dejana Zajc, Klemen Grabljevec
eSinapsa, 2014-7
Možgani v mreži navezanosti, ki nas zaznamuje
Barbara Horvat
Vpliv senzoričnega dotoka na uglasitev možganskih povezav
Peter Gradišnik
Človeški konektom ali kakšne so zveze v naših možganih
Blaž Koritnik
Niko Lah
Torkove delavnice za osnovnošolce
Mateja Drolec Novak, Vid V. Vodušek
Da ne pozabim! Tehnike za pomladitev spomina
Klara Tostovršnik, Hana Hawlina
Površina socialne nevroznanosti
Manuel Kuran
Clarity - bistri možgani Karla Deisserotha
Gregor Belušič
Barbara Gnidovec Stražišar
Bojana Žvan
Nevroplastičnost po možganski kapi
Marjan Zaletel
Klinično psihološka obravnava pacientov po možganski kapi in podpora pri vračanju na delovno mesto
Barbara Starovasnik Žagavec
Možgani: organ, s katerim ljubimo
Andraž Matkovič
Marija Šoštarič Podlesnik
Gibalno-kognitivna vadba: praktična delavnica
Mitja Gerževič, Marina Dobnik
Anton Grad
Nevrologija, imunologija, psihiatrija …
Bojan Rojc
Andraž Stožer, Janez Bregant
Dominika Novak Pihler
Možganska kap – »kako ostati v omrežju?«
Nina Ozimic
Klara Tostovršnik
eSinapsa, 2014-8
Znotrajžilno zdravljenje možganskih anevrizem
Tamara Gorjanc, Dimitrij Lovrič
Obravnava hladnih možganskih anevrizem
Bojana Žvan, Janja Pretnar Oblak
Ali deklice z Rettovim sindromom govorijo z očmi?
Anka Slana, Urška Slana
Progresivna multifokalna encefalopatija
Urša Zabret, Katarina Šurlan Popovič
Ne ubijaj – poskusi na živalih
Martina Perše
Poizkusi na živalih - za in proti
Simon Horvat
eSinapsa, 2015-9
Kako deluje navigacijski sistem v naših možganih
Simon Brezovar
Vsakodnevno delo slepe osebe / s slepo osebo
Denis Kamnar
Uroš Marušič
Manca Tekavčič Pompe
Toni Pustovrh
Marko Hawlina
Od svetlobe do podobe ali kako vidijo svet naši možgani
Simon Brezovar
Janja Hrastovšek
Zala Kurinčič
Pogledi na mejno osebnostno motnjo
Jerica Radež, Peter Kapš
Uvid kot socialno psihološki fenomen
Vid Vodušek
Uvod v vidno-prostorske funkcije s praktičnimi primeri
Ana Bujišić, Sanja Roškar
eSinapsa, 2015-10
Difuzijsko magnetnoresonančno slikanje
Rok Berlot
Katja Pavšič
Radiološko izolirani sindrom - ali ga moramo poznati?
Matej Vouk, Katarina Šurlan Popovič
Kako izgledajo možgani, ki govorijo več jezikov?
Gašper Zupan
Nov pristop v rehabilitaciji - terapija s pomočjo psa
Mateja Drljepan
Pogled v maternico z magnetnoresonančno preiskavo
Taja Jordan, Tina Vipotnik Vesnaver
Saša Zorjan
Saša Zorjan
Nevroestetika: ko nevroznanost obišče galerijo
Anja Voljavec, Hana Hawlina, Nika Vrabič
Ali so psihogeni neepileptični napadi res psihogeni?
Saška Vipotnik, Gal Granda
Kako nam lahko glasna glasba »vzame« sluh in povzroči tinitus
Nejc Steiner, Saba Battelino
eSinapsa, 2016-11
Mara Bresjanac
Kako ultrazvok odpira pot v možgane
Kaja Kolmančič
Kako je epigenetika spremenila nevroznanost
Metka Ravnik Glavač
Ondinino prekletstvo ali sindrom prirojene centralne hipoventilacije
Katja Pavšič, Barbara Gnidovec Stražišar, Janja Pretnar Oblak, Fajko F. Bajrović
Zika virus in magnetnoresonančna diagnostika nepravilnosti osrednjega živčevja pri plodu
Rok Banko, Tina Vipotnik Vesnaver
Motnje ravnotežja otrok in odraslih
Nejc Steiner, Saba Battelino
eSinapsa, 2016-12
Vloga magnetnoresonančne spektroskopije pri obravnavi možganskih tumorjev
Gašper Zupan, Katarina Šurlan Popovič
Tiskanje tridimenzionalnih modelov v medicini
Andrej Vovk
Aleš Oblak
Kevin Klarič
Sinestezija: umetnica, ki ne želi odrasti
Tisa Frelih
Računska psihiatrija: od nevroznanosti do klinike
Nastja Tomat
Kognitivni nadzor: od vsakdanjega življenja do bolezni
Vida Ana Politakis
eSinapsa, 2017-13
Internet: nadgradnja ali nadomestek uma?
Matej Perovnik
Vloga črevesnega mikrobioma pri odzivu na stres
Vesna van Midden
Stres pušča posledice tako na človeškem kot živalskem organizmu
Jasmina Kerčmar
Prikaz normalne anatomije in bolezenskih stanj obraznega živca z magnetno resonanco
Rok Banko, Matej Vrabec
Psihedelična izkušnja in njen zdravilni potencial
Anja Cehnar, Jona Basle
Vpliv hiperglikemije na delovanje možganov
Jasna Šuput Omladič, Simona Klemenčič
Nevrofibromatoza: napredujoče obolenje centralnega in perifernega živčevja
Nejc Steiner, Saba Battelino
Fenomen žrtvenega jagnja v dobi interneta
Dolores Trol
Tesnoba staršev in strategije spoprijemanja, ko pri otroku na novo odkrijejo epilepsijo
Daša Kocjančič, Petra Lešnik Musek, Vesna Krkoč, David Gosar
eSinapsa, 2017-14
Zakaj ne zapeljem s ceste, ko kihnem?
Anka Slana Ozimič, Grega Repovš
Nobelova nagrada za odkritje molekularnih mehanizmov nadzora cirkadianih ritmov
Leja Dolenc Grošelj
Možgani pod stresom: od celic do duševnih motenj
Nastja Tomat
Na sledi prvi vzročni terapiji Huntingtonove bolezni
Danaja Metul
Razlike med spoloma pri Parkinsonovi bolezni
Kaja Kolmančič
eSinapsa, 2018-15
Susceptibilno poudarjeno magnetnoresonančno slikanje pri bolniku z ALS
Alja Vičič, Jernej Avsenik, Rok Berlot
Sara Fabjan
Reverzibilni cerebralni vazokonstrikcijski sindrom – pot do diagnoze
Maja Cimperšek, Katarina Šurlan Popovič
Liam Korošec Hudnik
Kognitivno funkcioniranje pri izgorelosti
Marina Horvat
eSinapsa, 2019-16
Maša Čater
Saša Koprivec
Infekcije osrednjega živčnega sistema s flavivirusi
Maja Potokar
Raziskava: Kako depresija vpliva na kognitivne sposobnosti?
Vida Ana Politakis
Razvoj depresije pri otrocih z vidika navezovalnega vedenja
Neža Grgurevič
Sonja Prpar Mihevc
Umetno inteligentna nevroznanost: srečanje nevronskih mrež in možganske fiziologije
Kristijan Armeni
Čebelji strup pri preventivi nevrodegenerativnih bolezni in priložnost za klinično prakso
Matjaž Deželak
eSinapsa, 2019-17
IgG4+ – skupni imenovalec diagnoz iz preteklosti
Cene Jerele, Katarina Šurlan Popovič
Nov molekulski mehanizem delovanja ketamina v astrocitih
Matjaž Stenovec
Praktični pristop k obravnavi utrujenosti in motenj spanja pri bolnikih z multiplo sklerozo
Nik Krajnc, Leja Dolenc Grošelj
Jure Pešak
eSinapsa, 2020-18
Bolezni spektra anti-MOG pri odraslih
Nik Krajnc
Samomor pod lupo nevroznanosti
Alina Holnthaner
eSinapsa, 2020-19
Ob mednarodnem dnevu znakovnih jezikov
Anka Slana Ozimič
Teorija obetov: kako sprejemamo tvegane odločitve
Nastja Tomat
Sara Fabjan
Matjaž Deželak
Nina Stanojević, Uroš Kovačič
Od človeških nevronov do možganskih organoidov – nova obzorja v nevroznanosti
Vesna M. van Midden
Splošna umetna inteligenca ali statistične jezikovne papige?
Kristijan Armeni
Zunajcelični vezikli kot prenašalci zdravilnih učinkovin preko krvno-možganske prepreke
Saša Koprivec
Matjaž Deželak
eSinapsa, 2021-20
Migrena: starodavna bolezen, sodobni pristopi k zdravljenju
Eva Koban, Lina Savšek
Zgodnji razvoj socialnega vedenja
Vesna Jug
Nastja Tomat
Mikrosplet: povezovanje preko mikrobioma
Tina Tinkara Peternelj
Stimulacija možganov kot način zdravljenja depresije
Saša Kocijančič Azzaoui
eSinapsa, 2021-21
eSinapsa, 2022-22
Sodobni vidiki motenj hranjenja
Karin Sernec
Ples in gibalni dialog z malčki
Neva Kralj
Atul Gawande
Jezikovna funkcija pri Alzheimerjevi bolezni
Gašper Tonin
Dostava terapevtikov preko krvno-možganske pregrade
Matjaž Deželak
eSinapsa, 2022-23
Akutni ishemični infarkt hrbtenjače pri zdravih otrocih – kaj lahko pove radiolog?
Katarina Šurlan Popovič, Barbara Šijaković
eSinapsa, 2023-24
Možganska omrežja pri nevrodegenerativnih boleznih
Tomaž Rus, Matej Perovnik
Morske živali kot navdih za nevroznanstvenike: morski konjiček, morski zajček in klobučnjak
Tina Bregant
Metoda Feldenkrais: gibanje in nevroplastičnost
Mateja Pate
Etično naravnana animalna nevroznanost
Maša Čater
Helena Motaln, Boris Rogelj
eSinapsa, 2023-25
Urban Košak, Damijan Knez, Anže Meden, Simon Žakelj, Jurij Trontelj, Jure Stojan, Maja Zakošek Pipan, Kinga Sałat idr.
eSinapsa, 2024-26
Naravno okolje kot vir zdravja in blagostanja
Karin Križman, Grega Repovš, Gaja Zager Kocjan, Gregor Geršak
Katja Peganc Nunčič, Damjan Osredkar
Tanja Goltnik
Ali je zgodnje vstajanje dedno?
Cene Skubic, Laura Plavc, Damjana Rozman, Leja Dolenc Grošelj
eSinapsa, 2024-27
Širša terapevtska uporaba ketamina: potenciali in izzivi
Kristian Elersič
Moč vpliva socialne opore na bolečino
Jana Verdnik
Benjamin Bušelič
Urška Černe, Anemari Horvat, Robert Zorec, Nina Vardjan
eSinapsa, 2025-28
Maša Čater, Nastja Blagovič, Urška Jerič, Agata Kokalj Malovrh, Nuša Balen, Tanja Kunej
Vpliv izobrazbe na skrb za zdravje možganov
Hana Kos, Matej Perovnik
Selena Horvat, Anja Pišlar
eSinapsa, 2025-29
Knjiga Kje so moji ključi kot primer narativne medicine
Zdenka Čebašek-Travnik, Saša Novak
Vpliv anestetikov na oksidativni stres
Katerina Tomsič, Alenka Nemec Svete
Rak, živčni in imunski sistem – kako so med seboj povezani?
Maja Čemažar, Tanja Jesenko, Urša Lampreht Tratar, Maša Omerzel
eSinapsa, 2026-30
Denis Štepihar, Klementina Fon Tacer
Marta Macedoni Lukšič
eSinapsa, 2026-31
Otroci se v prvem razredu osnove šole učijo pisanja in prepoznavanja črk, nadaljujejo pa s povezovanjem le-teh v besede in nato več besed v stavke. Spoznavanje s črkami in zanimanje zanje pa se po navadi prične že mnogo prej. Prav vznemirljivo je, kako hitro otroci naredijo preskok v znanju. Samo nekaj tednov vaje je potrebnih in že se iz popolnega nebralca prelevijo v bralca. S časom se veščina branja še utrdi in izboljša. Ob učenju branja pa poteka kompleksen razvoj možganov.
Razvoj pismenosti
Branje in pisanje med drugim vključujeta mentalno predstavo nabora vidnih simbolov, ki so povezani v besede preko različnih fonoloških pravil. Potrebno je tekoče povezovanje med semantičnim besediščem, ki predstavlja shrambo pomena besed, fonološkimi predstavami, ki velja za shrambo slušnega zvoka besede, ter ortografskim besediščem, ki združuje znanje vidne podobe napisanih besed. Ti trije tipi besedišča niso podedovani, temveč se razvijejo kot posledica izpostavljenosti materinemu jeziku. Tako možgani niso genetsko pogojeni s samim razvojem pismenosti, ampak se z izpostavljenostjo vidnim in slušnim jezikovnim dražljajem učijo brati in pisati, kar pa zahteva reorganizacijo možganov 1.
Spremembe v možganih kot podlaga za razvoj bralnih sposobnosti
Branje velja za priučeno veščino, ki je močno odvisna predvsem od dveh parametrov; od razvitosti možganov ter bralnih izkušenj. Razvoj možganov poteka na podlagi sprememb v sivini in belini možganov. Bela možganovina sestavlja aksonske povezave v notranjosti možganov, medtem ko skorjo in možganska jedra v notranjosti poimenujemo kot sivino. Spremembe ob razvoju možganov so tako regionalno kot časovno specifične; v splošnem velja, da se volumen sivine zmanjšuje, medtem ko se volumen beline povečuje med prehodom iz otroštva v odraslost. V filogenetsko starejših regijah možganov pride do sprememb v volumnu možganovine pri razvoju možganov prej kot v filogenetsko novejših regijah. Spremembe v sivini in belini so pod vplivom tako genetike kot okolja. Genetika vpliva na razvoj delov možganov v različni meri. Pri otrocih in mladostnikih so ugotovili, da genetski faktorji v največji meri (80 %) vplivajo na celotni volumen možganov in v podobni meri (77% in 89 %) tudi posamično na volumen sivine in beline. Razlike v vplivu so celo med hemisferama, saj je volumen desne hemisfere bolj podvržen vplivu genov (v deležu 66 %) kot volumen leve (56 %) 2. Po drugi strani tudi izkušnje iz okolja močno vplivajo na razvoj možganov pri otrocih. Psihosocialni dejavniki iz okolja vplivajo na razvoj nevronov in na spremembe v določenih delih možganov. Potrjen je na primer vpliv materine podpore in bližine v predšolski dobi na hitrejšo rast hipokampusa pri otrocih v šolski dobi in kasneje v odraslosti. Bolj razvit hipokampus pa se nato odraža v bolje razvitem čustvenem razvoju, spominu in sposobnosti uravnavanja stresa 3. Z ozirom na razvoj sposobnosti branja lahko iz najnovejših dognanj povzamemo, da pri otrocih, ki so boljši bralci in ki berejo hitreje ter pogosteje kot nekoliko slabši bralci, prihaja do drugačnega razvoja določenih regij v možganih 4.
Z branjem povezani deli možganov
Z metodami slikanja možganov so dokazali, da branje omogoča in izvaja področje leve hemisfere, kjer pride do mreženja nevronov iz frontalnih, temporo-parietalnih in okcipito-temporalnih predelov skorje (slika 1). Ti omogočajo povezavo med vidnimi (ortografskimi) in slušnimi (fonološkimi) informacijami ter konceptualnimi (semantičnimi) podobami. Otroški možgani hitro vzpostavljajo nove sinaptične povezave, kar omogoča nove miselne sposobnosti. Pri tem prihaja do večje in natančnejše funkcionalne in strukturne povezanosti regij možganov med seboj. Slušni korteks, odgovoren za sprejem glasovnih predstav, se povezuje s predeli za procesiranje glasov – levi inferiorni frontalni girus 1.
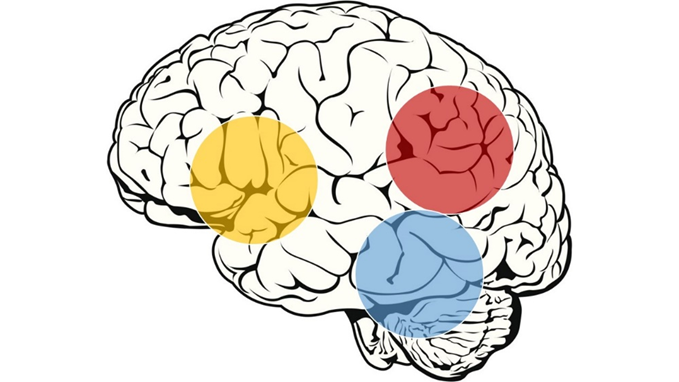 Slika 1: Anatomski deli možganske skorje leve hemisfere, ki so povezani z branjem. Rumena – inferio-frontalni del, rdeča – temporo-parietalni del, modra – okcipito-temporalni del.
Slika 1: Anatomski deli možganske skorje leve hemisfere, ki so povezani z branjem. Rumena – inferio-frontalni del, rdeča – temporo-parietalni del, modra – okcipito-temporalni del.
Disleksija in aleksija
Ena izmed motenj v učenju branja je nevrobiološka motnja disleksija. Tako pri otrocih kot odraslih so z nevrološkim slikanjem dokazali zmanjšano aktivnost levega temporo-parietalnega in okcipito-temporalnega predela skorje med fonološkim procesiranjem in branjem. Disleksični bralci pravzaprav izkazujejo povečano aktivnost desne hemisfere in bilateralnih prefrontalnih dorzalnih predelov, kar naj bi predstavljalo kompenzacijo za slabše delovanje levih posteriornih predelov 1. Disleksija velja za dedno nevrološko razvojno motnjo, ki se izraža kot konstantno, doživljenjsko oteženo branje in/ali črkovanje, ki pa ni povezana z nižjo inteligenco ali okoljskimi vplivi 5. Motnje branja se lahko kažejo na različne načine. Dislektiki pogosto težko prepoznavajo besede, ki jih sicer že poznajo, in jih težko črkujejo. Težave imajo pri pisanju, hitrem branju, izgovarjavi besed ter pri razumevanju pisane besede. Pogosto imajo disleksiki težave tudi s številkami. Dednost igra pomembno vlogo pri pojavnosti disleksije. Le-ta se pojavi pri otrocih, kjer je eden od staršev disleksičen, kar v 40-60 %. V kolikor sta disleksična oba starša, pa se ta delež še precej poveča 6. Pri otrocih disleksičnih staršev so odzivi možganov drugačni že od rojstva naprej. Raziskave s funkcionalno magnetno resonanco so pokazale, da je pri disleksikih funkcijsko povezovanje regij možganov, ki sodelujejo pri branju, precej slabše 7. Drugačni odzivi možganov pri disleksiji so povezani s spremembami v anatomiji primarnega slušnega korteksa oziroma superiotemporalnega korteksa in tudi drugih okcipito-temporalnih, posteriotemporalnih in okcipitalnih regij.
Novejše študije potrjujejo povezavo med tekočim branjem in spremembo v volumnu možganske skorje pri otrocih v razvoju 8, 9. Raziskovalci so ugotovili, da je z učinkovitim branjem povezan upad volumna frontalnega in parietalnega režnja možganske skorje 4. Na podlagi merjenja odzivov možganov pri dojenčku je celo možno napovedati hitrost branja, ki jo bo usvojil v obdobju adolescence 10. Pridobljena nezmožnost branja, aleksija, pa se lahko pojavi pri ljudeh, ki so že brali, vendar so izgubili to sposobnost, bodisi po kapi ali poškodbi področja v možganih, ki je povezano z bralnimi sposobnostmi. Najpogosteje je aleksija posledica infarkta leve posteriorne cerebralne arterije, kar se kaže v tem, da oseba lahko piše, ne more pa brati 11, 12.
V splošnem torej velja, da je intaktna leva hemisfera možganov, še posebno temporoparietalni del, izrednega pomena za dobro bralno sposobnost. Vendar kompleksnost razvoja procesa pridobivanja sposobnosti branja ne temelji le na aktivaciji ustreznih delov skorje, temveč tudi in predvsem na ustrezni komunikaciji med njimi. Raziskave o natančni identifikaciji povezav med anatomijo in funkcijo delov možganov v povezavi z branjem se nadaljujejo in nam bodo v prihodnosti omogočile natančnejši vpogled v dejansko shemo procesa učenja branja.
-
___
-
Vandermosten M., Boets B., Wouters J., Ghesquiere P. A qualitative and quantitative review of diffusion tensor imaging studies in reading and dyslexia. Neuroscience and Biobehavioral Reviews. 2012;36(6):1532-1552. ↩
-
Kremen W. S., Prom-Wormley E., Panizzon M. S., Eyler L. T., Fischl B., Neale M. C., Franc C. E., Lyons M. J., Pacheco J., Perry M. E., Stevens A., Schmitt J. E., Grant M. D., Sediman L. J., Thermenos H. W., Tsuang M. T., Eisen S. A., Dale A. M. Fennema-Notestine C. Genetic and environmental influences on the size of specific brain regions in midlife: the VETSA MRI study. Neuroimage. 2010;49(1):1213-1223. ↩
-
Luby J. L., Barch D. M., Belden A., Gaffrey M. S., Tillman R., Babb C., Nishino T., Suzuki H., Botteron K. N. Maternal support in early childhood predicts larger hippocampal volumes at school age. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(8):2854-2859. ↩
-
Houston S. M., Lebel C., Katzir T., Manis F. R., Kan E., Rodriguez G. R., Sowell E. R. Reading skill and structural brain development. Neuroreport. 2014;25(5):347-352. ↩
-
Vellutino F.R., Fletcher J.M., Snowling M.J., Scanlon D.M. Specific reading disability (dyslexia): what have we learned in the past four decades? Journal of Child Psychology and Psychiatry. 2004;45:2–40. ↩
-
Schumacher J., Hoffmann P., Schmal C., Schulte-Korne G., Nothen M. M. Genetics of dyslexia: the evolving landscape. Journal of Medical Genetics. 2007;44(5):289-297. ↩
-
Boets B., Op de Beeck H. P., Vandermosten M., Scott S. K., Gillebert C. R., Mantini D., Bulthe J., Sunaert S., Wouters J., Ghesquiere P. Intact but less accessible phonetic representations in adults with dyslexia. Science. 2013;342(6163):1251-1254. ↩
-
Sowell E. R., Thompson P. M., Leonard C. M., Welcome S. E., Kan E., Toga A. W. Longitudinal mapping of cortical thickness and brain growth in normal children. Journal of Neuroscience. 2004;24:8223–8231. ↩
-
Squeglia L. M., Jacobus J., Sorg S. F., Jerniga T. L., Tapert ,S. F. Early adolescentcortical thinning is relatedto better neuropsychological performance. Journal of the International Neuropsychological Society. 2013;19:962–970. ↩
-
Lohvansuu K., Hamalainen J. A., Ervast L., Lyytinen H., Leppanen P. H. T. Longitudinal interactions between brain and cognitive measures on reading development from 6 months to 14 years. Neuropsychologia. 2018;108:6-12. ↩
-
Kim E. S., Rising K., Rapcsak S. Z., Beeson P. M. Treatment for Alexia With Agraphia Following Left Ventral Occipito-Temporal Damage: Strengthening Orthographic Representations Common to Reading and Spelling. JSLHR. 2015;58:1521. ↩
-
Ožura A., Horvat Ledinek A., Starovasnik B., Sever A., Vodušek D. B. Aleksija brez agrafije pri tumorski obliki multiple skleroze: prikaz primera. Zdravstveni vestnik. 2009;78:9-12. ↩
Asist. – razisk. dr. Maša Čater, univ. dipl. bioteh.
Laboratorij za genomiko
Inštitut za predklinične vede
Veterinarska fakulteta
Univerza v Ljubljani
Sprejeto: 17.12.2018
Objavljeno: 14.1.2019