Spletna revija za znanstvenike, strokovnjake
in nevroznanstvene navdušence
Naslovnica Članki Intervjuji Mnenja Zdravje Korenine eSinapsa Številke ![]()
Je skrb za potomce prirojena, privzgojena ali pridobljena?
članki
eSinapsa, 2011-1
Zvezdan Pirtošek
Eksoskeleti – inteligentne bionske naprave
Marko Munih
O aktualnih dilemah draženja globokih možganskih struktur pri obsesivno - kompulzivni motnji
Nadja Jarc
Sledite svojo srečo ... z iPhone
Urban Kordeš
eSinapsa, 2011-2
Renata Salecl
Gašper Tkačik
Astrociti – spregledane zvezde nevrobiologije
Marko Kreft, Robert Zorec
Sašo Dolenc
Meditacija - malo truda, veliko koristi
Luka Dimic
eSinapsa, 2011-3
Mara Bresjanac
Martina Starc
Rok Berlot
Varnost uporabe generičnih protiepileptičnih zdravil
Mojca Kržan, Matevž Kržan
Možgani, računalniki - nekaj vmes
Miha Pelko
eSinapsa, 2012-4
Ali so moški in ženski možgani različni?
Gregor Majdič
O kognitivnih motnjah pri bolnikih s Parkinsonovo boleznijo
Dejan Georgiev
Akutno možgansko kap lahko uspešno zdravimo
Nina Vujasinovič, Bojana Žvan
Vloga nevropsihološke diagnostike pri odkrivanju zgodnjih znakov alzheimerjeve bolezni
Simon Brezovar
eSinapsa, 2013-5
Subarahnoidna krvavitev zaradi tromboze venskih sinusov
Mateja Repar, Anita Resman Gašperčič
Srečanje dveh velikanov: možganov in imunskega sistema
Matej Markota
Novo odkritje na področju sporadičnih prionskih bolezni
Jana Jerše, Nadja Jarc
Učinek placeba brez lažnih zdravil in zavajanja
Mara Bresjanac
eSinapsa, 2013-6
Odstranjevanje možganskih tumorjev pri budnem bolniku
Andrej Vranič, Jasmina Markovič, Blaž Koritnik
Zmedena bolnica, ki nič ne vidi ali PRES
Manja Hribar, Vid Zgonc
Manja Hribar
Sistemska skleroza in ishemična možganska kap - vzročna povezanost ali le koincidenca?
Mateja Repar, Janja Pretnar Oblak
Netravmatska lokalizirana konveksitetna subarahnoidna krvavitev
Mateja Repar, Fajko F. Bajrović
Klemen Grabljevec
Z omejevanjem spodbujajoča terapija pri bolnikih po nezgodni možganski poškodbi
Dejana Zajc, Klemen Grabljevec
eSinapsa, 2014-7
Možgani v mreži navezanosti, ki nas zaznamuje
Barbara Horvat
Vpliv senzoričnega dotoka na uglasitev možganskih povezav
Peter Gradišnik
Človeški konektom ali kakšne so zveze v naših možganih
Blaž Koritnik
Niko Lah
Torkove delavnice za osnovnošolce
Mateja Drolec Novak, Vid V. Vodušek
Da ne pozabim! Tehnike za pomladitev spomina
Klara Tostovršnik, Hana Hawlina
Površina socialne nevroznanosti
Manuel Kuran
Clarity - bistri možgani Karla Deisserotha
Gregor Belušič
Barbara Gnidovec Stražišar
Bojana Žvan
Nevroplastičnost po možganski kapi
Marjan Zaletel
Klinično psihološka obravnava pacientov po možganski kapi in podpora pri vračanju na delovno mesto
Barbara Starovasnik Žagavec
Možgani: organ, s katerim ljubimo
Andraž Matkovič
Marija Šoštarič Podlesnik
Gibalno-kognitivna vadba: praktična delavnica
Mitja Gerževič, Marina Dobnik
Anton Grad
Nevrologija, imunologija, psihiatrija …
Bojan Rojc
Andraž Stožer, Janez Bregant
Dominika Novak Pihler
Možganska kap – »kako ostati v omrežju?«
Nina Ozimic
Klara Tostovršnik
eSinapsa, 2014-8
Znotrajžilno zdravljenje možganskih anevrizem
Tamara Gorjanc, Dimitrij Lovrič
Obravnava hladnih možganskih anevrizem
Bojana Žvan, Janja Pretnar Oblak
Ali deklice z Rettovim sindromom govorijo z očmi?
Anka Slana, Urška Slana
Progresivna multifokalna encefalopatija
Urša Zabret, Katarina Šurlan Popovič
Ne ubijaj – poskusi na živalih
Martina Perše
Poizkusi na živalih - za in proti
Simon Horvat
eSinapsa, 2015-9
Kako deluje navigacijski sistem v naših možganih
Simon Brezovar
Vsakodnevno delo slepe osebe / s slepo osebo
Denis Kamnar
Uroš Marušič
Manca Tekavčič Pompe
Toni Pustovrh
Marko Hawlina
Od svetlobe do podobe ali kako vidijo svet naši možgani
Simon Brezovar
Janja Hrastovšek
Zala Kurinčič
Pogledi na mejno osebnostno motnjo
Jerica Radež, Peter Kapš
Uvid kot socialno psihološki fenomen
Vid Vodušek
Uvod v vidno-prostorske funkcije s praktičnimi primeri
Ana Bujišić, Sanja Roškar
eSinapsa, 2015-10
Difuzijsko magnetnoresonančno slikanje
Rok Berlot
Katja Pavšič
Radiološko izolirani sindrom - ali ga moramo poznati?
Matej Vouk, Katarina Šurlan Popovič
Kako izgledajo možgani, ki govorijo več jezikov?
Gašper Zupan
Nov pristop v rehabilitaciji - terapija s pomočjo psa
Mateja Drljepan
Pogled v maternico z magnetnoresonančno preiskavo
Taja Jordan, Tina Vipotnik Vesnaver
Saša Zorjan
Saša Zorjan
Nevroestetika: ko nevroznanost obišče galerijo
Anja Voljavec, Hana Hawlina, Nika Vrabič
Ali so psihogeni neepileptični napadi res psihogeni?
Saška Vipotnik, Gal Granda
Kako nam lahko glasna glasba »vzame« sluh in povzroči tinitus
Nejc Steiner, Saba Battelino
eSinapsa, 2016-11
Mara Bresjanac
Kako ultrazvok odpira pot v možgane
Kaja Kolmančič
Kako je epigenetika spremenila nevroznanost
Metka Ravnik Glavač
Ondinino prekletstvo ali sindrom prirojene centralne hipoventilacije
Katja Pavšič, Barbara Gnidovec Stražišar, Janja Pretnar Oblak, Fajko F. Bajrović
Motnje ravnotežja otrok in odraslih
Nejc Steiner, Saba Battelino
Zika virus in magnetnoresonančna diagnostika nepravilnosti osrednjega živčevja pri plodu
Rok Banko, Tina Vipotnik Vesnaver
eSinapsa, 2016-12
Vloga magnetnoresonančne spektroskopije pri obravnavi možganskih tumorjev
Gašper Zupan, Katarina Šurlan Popovič
Tiskanje tridimenzionalnih modelov v medicini
Andrej Vovk
Aleš Oblak
Kevin Klarič
Sinestezija: umetnica, ki ne želi odrasti
Tisa Frelih
Računska psihiatrija: od nevroznanosti do klinike
Nastja Tomat
Kognitivni nadzor: od vsakdanjega življenja do bolezni
Vida Ana Politakis
eSinapsa, 2017-13
Internet: nadgradnja ali nadomestek uma?
Matej Perovnik
Vloga črevesnega mikrobioma pri odzivu na stres
Vesna van Midden
Stres pušča posledice tako na človeškem kot živalskem organizmu
Jasmina Kerčmar
Prikaz normalne anatomije in bolezenskih stanj obraznega živca z magnetno resonanco
Rok Banko, Matej Vrabec
Psihedelična izkušnja in njen zdravilni potencial
Anja Cehnar, Jona Basle
Vpliv hiperglikemije na delovanje možganov
Jasna Šuput Omladič, Simona Klemenčič
Nevrofibromatoza: napredujoče obolenje centralnega in perifernega živčevja
Nejc Steiner, Saba Battelino
Fenomen žrtvenega jagnja v dobi interneta
Dolores Trol
Tesnoba staršev in strategije spoprijemanja, ko pri otroku na novo odkrijejo epilepsijo
Daša Kocjančič, Petra Lešnik Musek, Vesna Krkoč, David Gosar
eSinapsa, 2017-14
Zakaj ne zapeljem s ceste, ko kihnem?
Anka Slana Ozimič, Grega Repovš
Nobelova nagrada za odkritje molekularnih mehanizmov nadzora cirkadianih ritmov
Leja Dolenc Grošelj
Možgani pod stresom: od celic do duševnih motenj
Nastja Tomat
Na sledi prvi vzročni terapiji Huntingtonove bolezni
Danaja Metul
Razlike med spoloma pri Parkinsonovi bolezni
Kaja Kolmančič
eSinapsa, 2018-15
Susceptibilno poudarjeno magnetnoresonančno slikanje pri bolniku z ALS
Alja Vičič, Jernej Avsenik, Rok Berlot
Sara Fabjan
Reverzibilni cerebralni vazokonstrikcijski sindrom – pot do diagnoze
Maja Cimperšek, Katarina Šurlan Popovič
Liam Korošec Hudnik
Kognitivno funkcioniranje pri izgorelosti
Marina Horvat
eSinapsa, 2019-16
Maša Čater
Saša Koprivec
Infekcije osrednjega živčnega sistema s flavivirusi
Maja Potokar
Raziskava: Kako depresija vpliva na kognitivne sposobnosti?
Vida Ana Politakis
Razvoj depresije pri otrocih z vidika navezovalnega vedenja
Neža Grgurevič
Sonja Prpar Mihevc
Umetno inteligentna nevroznanost: srečanje nevronskih mrež in možganske fiziologije
Kristijan Armeni
Čebelji strup pri preventivi nevrodegenerativnih bolezni in priložnost za klinično prakso
Matjaž Deželak
eSinapsa, 2019-17
IgG4+ – skupni imenovalec diagnoz iz preteklosti
Cene Jerele, Katarina Šurlan Popovič
Nov molekulski mehanizem delovanja ketamina v astrocitih
Matjaž Stenovec
Praktični pristop k obravnavi utrujenosti in motenj spanja pri bolnikih z multiplo sklerozo
Nik Krajnc, Leja Dolenc Grošelj
Jure Pešak
eSinapsa, 2020-18
Bolezni spektra anti-MOG pri odraslih
Nik Krajnc
Samomor pod lupo nevroznanosti
Alina Holnthaner
eSinapsa, 2020-19
Ob mednarodnem dnevu znakovnih jezikov
Anka Slana Ozimič
Teorija obetov: kako sprejemamo tvegane odločitve
Nastja Tomat
Sara Fabjan
Matjaž Deželak
Nina Stanojević, Uroš Kovačič
Od človeških nevronov do možganskih organoidov – nova obzorja v nevroznanosti
Vesna M. van Midden
Splošna umetna inteligenca ali statistične jezikovne papige?
Kristijan Armeni
Zunajcelični vezikli kot prenašalci zdravilnih učinkovin preko krvno-možganske prepreke
Saša Koprivec
Matjaž Deželak
eSinapsa, 2021-20
Migrena: starodavna bolezen, sodobni pristopi k zdravljenju
Eva Koban, Lina Savšek
Zgodnji razvoj socialnega vedenja
Vesna Jug
Nastja Tomat
Mikrosplet: povezovanje preko mikrobioma
Tina Tinkara Peternelj
Stimulacija možganov kot način zdravljenja depresije
Saša Kocijančič Azzaoui
eSinapsa, 2021-21
eSinapsa, 2022-22
Sodobni vidiki motenj hranjenja
Karin Sernec
Ples in gibalni dialog z malčki
Neva Kralj
Atul Gawande
Jezikovna funkcija pri Alzheimerjevi bolezni
Gašper Tonin
Dostava terapevtikov preko krvno-možganske pregrade
Matjaž Deželak
eSinapsa, 2022-23
Akutni ishemični infarkt hrbtenjače pri zdravih otrocih – kaj lahko pove radiolog?
Katarina Šurlan Popovič, Barbara Šijaković
eSinapsa, 2023-24
Možganska omrežja pri nevrodegenerativnih boleznih
Tomaž Rus, Matej Perovnik
Morske živali kot navdih za nevroznanstvenike: morski konjiček, morski zajček in klobučnjak
Tina Bregant
Metoda Feldenkrais: gibanje in nevroplastičnost
Mateja Pate
Etično naravnana animalna nevroznanost
Maša Čater
Helena Motaln, Boris Rogelj
eSinapsa, 2023-25
Urban Košak, Damijan Knez, Anže Meden, Simon Žakelj, Jurij Trontelj, Jure Stojan, Maja Zakošek Pipan, Kinga Sałat idr.
eSinapsa, 2024-26
Naravno okolje kot vir zdravja in blagostanja
Karin Križman, Grega Repovš, Gaja Zager Kocjan, Gregor Geršak
Katja Peganc Nunčič, Damjan Osredkar
Tanja Goltnik
Ali je zgodnje vstajanje dedno?
Cene Skubic, Laura Plavc, Damjana Rozman, Leja Dolenc Grošelj
eSinapsa, 2024-27
Širša terapevtska uporaba ketamina: potenciali in izzivi
Kristian Elersič
Moč vpliva socialne opore na bolečino
Jana Verdnik
Benjamin Bušelič
Urška Černe, Anemari Horvat, Robert Zorec, Nina Vardjan
eSinapsa, 2025-28
Maša Čater, Nastja Blagovič, Urška Jerič, Agata Kokalj Malovrh, Nuša Balen, Tanja Kunej
Vpliv izobrazbe na skrb za zdravje možganov
Hana Kos, Matej Perovnik
Selena Horvat, Anja Pišlar
eSinapsa, 2025-29
Knjiga Kje so moji ključi kot primer narativne medicine
Zdenka Čebašek-Travnik, Saša Novak
Vpliv anestetikov na oksidativni stres
Katerina Tomsič, Alenka Nemec Svete
Rak, živčni in imunski sistem – kako so med seboj povezani?
Maja Čemažar, Tanja Jesenko, Urša Lampreht Tratar, Maša Omerzel
eSinapsa, 2026-30
Denis Štepihar, Klementina Fon Tacer
Marta Macedoni Lukšič
eSinapsa, 2026-31
Starševska skrb se izraža na različne načine: od telesne skrbi in nege za potomce, vključno z dojenjem, do motivacije za druženje z njimi in pri ljudeh tudi skrbi za njihovo ekonomsko dobrobit. Kot socialno-vedenjsko strategijo lahko starševsko vedenje proučujemo z nevrološkega vidika. V prispevku predstavljamo možgansko regijo, ki je dosledno opredeljena kot bistvena pri skrbi za potomce. Gre za volumsko majhno področje v hipotalamusu, imenovano srednje predoptično območje (angl. medial preoptic nucleus, MPOA). Med drugim ga sestavlja posebna skupina nevronov (MPOAGal nevroni), ki izražajo nevropeptid galanin. Ti nevroni so prisotni pri obeh spolih in so ključni za starševsko obnašanje. Z razmišljanjem o nevroloških osnovah za starševsko vedenje se dotaknemo tudi dveh skrajnih aspektov starševstva: žrtvovanja lastne koristi v dobro potomcev na eni strani ter detomora na drugi. Z razumevanjem starševskega vedenja drugih sesalcev bomo tako morda bolje razumeli tudi lastno vedenje.
Uvod
Ljudje posvetimo vzgoji svojih potomcev praviloma dve do tri desetletja. V zadnjem času se kljub družbi izobilja, ali pa morda prav zato, skrb za potomce pri ljudeh celo podaljšuje. Ne gre le za materialno ali fizično preskrbljenost, pač pa tudi za čustveno in socialno skrb zanje. Lahko se vprašamo, kaj nas pravzaprav žene, da toliko časa, energije in denarja namenimo svojim potomcem?
Skrb za potomce
Skrb za potomce lahko opazujemo pri različnih živalskih vrstah in praktično pri vseh taksonomskih skupinah. Zanimiva je skrb za potomce pri socialnih žuželkah, kot so mravlje in čebele, opazimo jo pri ribah, pticah in seveda sesalcih, vključno pri nas, ljudeh. Gre za socialno-vedenjsko evolucijsko strategijo. Bolj ekonomsko bi lahko rekli, da gre za starševsko naložbo v razvojno sposobnost potomcev. Kljub povečani razvojni sposobnosti potomcev in posledično povečanju preživetvene sposobnosti celotne vrste, pa ima ta skrb svojo ceno. Starši izdatno trošijo energijo za potomce in skrb zanje ter tako »izgubljajo« oziroma »zapravljajo« lastne razvojne možnosti, predvsem vsaj začasno zanemarijo možnost razmnoževanja 1. Zato se je strategija skrbi za potomce ohranila le pri tistih vrstah, kjer so evolucijsko gledane koristi za potomce večje od »žrtvovanja« staršev 2. Strategija skrbi za potomce je odvisna tudi od okolja 3. Ob pomanjkanju hrane v okolju lahko opazimo, da se skrb za potomce zmanjša, saj zahteva precejšen vložek staršev. Od razpoložljivosti dobrin je odvisno tudi ravnanje staršev v podobnih okoliščinah: v obdobju obilja tako starši najbolj pogosto hranijo tistega mladiča, ki najbolj prosi za hrano, nasprotno pa v obdobju pomanjkanja, mladiča s podobnim vedenjem ignorirajo 4.
Starševsko vedenje pri drugih vrstah
Živali kažejo zelo raznolike oblike starševskega vedenja, ki temeljijo na socioekologiji, energetskih potrebah, omejitvah pri razmnoževanju (npr. številčnost potomcev) in načinu hranjenja potomcev (npr. dojenje pri sesalcih). Pri dvoživkah in plazilcih najdemo le malo komponent starševskega vedenja 5; pri več kot 80 % ptic najdemo starševsko vedenje, ki zahteva prisotnost obeh staršev 6. Strupene, barvite žabice Srednje in Južne Amerike, kot so podrevnice (družina Dendrobatidae), izkazujejo različna starševska vedenja, odvisno od selekcijskih pritiskov okolja. Večinoma izkazujejo izvorno enostransko, moško skrb za potomce; v zahtevnih razmerah, na primer v majhnih ribnikih, pa se poslužujejo energetsko zahtevnih strategij, ki vključujejo tako hranjenje paglavcev z neoplojenimi jajčeci kot tudi skrb za potomce s strani obeh staršev oz. ali samo matere 7.
Pri laboratorijskih živalih (miši, podgane, opice) smo pogosto omejeni na sesalce, ki izkazujejo prvenstveno enostarševsko nego, konkretno materino. Mnoge vrste sicer izkazujejo skupno skrb za potomce, ko sorodniki ali pripadniki iste skupine pomagajo skrbeti za potomce 8. Slednje naj bi veljalo zlasti za prednike ljudi, kar naj bi jih bistveno razlikovalo od ostalih primatov, ki so do svojih potomcev zelo posesivni. Deljena skrb za potomce, zlasti s strani babic, ki pomagajo tako svojim hčerkam kot vnukom, naj bi omogočala razvoj skupnosti, ki je odvisna eden od drugega in medsebojne pomoči, ter tudi razcvet človeštva.
Straševstvo pri ljudeh
Strategija starševstva in razmnoževanja je pri ljudeh zelo raznolika. Za današnje dni bizarno, lahko v Guinnessovi knjigi rekordov iz leta 1998 preberemo, da naj bi v 18. stoletju v Rusiji živel izjemno ploden par 9. Ruski kmet Feodor Vasiljev naj bi imel s prvo ženo kar 69 otrok, od tega 16 dvojčkov, 7 trojčkov in 4 četvorčke. Nato je Feodor imel še eno ženo in z njo 18 otrok 10. Nekoliko presenetljiva za današnje dni je genetska dediščina Džingiskana in njegovih moških sorodnikov, saj naj bi kar 8 % današnjih moških v Aziji izviralo iz njegove družine 11. Celo več, evolucijski demografi so izračunali, da je možno, da ima en moški s približno 65–110 ženskami številčno najvišji izplen potomcev – skoraj 1000 otrok 12.
Evolucijski demografi so navdih poiskali v zgodovinskih knjigah, kjer so razbrali, da naj bi maroški vladar Mulaj Ismael Krvoločni zaplodil kar 888 otrok 12. Je pa zanimivo, da so tudi izračunali, da se večji harem kot 100 žena bogatemu moškemu niti ne izplača z vidika potomstva, saj ne rezultira v toliko več potomcih. Ob teh zabavno bizarnih analizah demografi opozarjajo na pomembne družbene mehanizme, ki vplivajo tako na odločitev, kdaj imeti potomce in koliko; ekonomsko-finančno breme je pri teh odločitvah zelo pomembno 13.
Starševstvo in skrb za vzgojo sta pri ljudeh izjemno zahtevna. Potrebna je pozornost, ki povprečno zahteva 13 milijonov kalorij vložka do otrokovega 18. leta 14. Izračun je bil narejen na podlagi analize treh tradicionalnih lovsko-nabiralniških skupnosti Južne Amerike, vključuje pa tudi sposobnost odpovedovanju lastnim potrebam, od spanca do prostega časa, in to brez takojšnjega učinka. Svoj vložek in napor lahko starši ocenjujemo šele mnogo let kasneje, ko z vlažnimi očmi opazujemo svojega otroka na maturantskem plesu ali ob prejemu diplome. Po poročilu ZDA o izdatkih za otroke po družinah znaša skrb za otroka, rojenega v ZDA v štiričlanski družini s povprečnim dohodkom (ki pa je višji kot v Sloveniji!), leta 2013 do njegovega dopolnjenega 18. leta kar 304.480 USD oziroma 268.630 EUR in to brez stroškov šolanja in prilagojeno napovedani inflaciji 15. Tudi v Sloveniji poznamo okvirne stroške za vzgojo otroka od rojstva pa do konca srednje šole, ki znašajo glede na evropsko povprečje približno 100.000 EUR oziroma približno četrtino povprečnega družinskega proračuna 16. Pri izračunih gre le za okvirne vrednosti; ekonomsko-socialne razmere se tudi spreminjajo. Denar pa ni vse in pri skrbi za potomce izjemno štejejo stvari, ki se jih ne da finančno ovrednotiti: skrb, ljubezen, navezanost, občutek pripadnosti. Na ta občutja vplivajo naši možgani. Morda se tudi zato odgovor na neekonomičnost, a vendar evolucijsko stalnost starševskega vedenja, skriva prav v možganih?
Ali je starševstvo zapisano v možganih?
Skozi evolucijo so se izoblikovali in ohranili mehanizmi, ki omogočajo krepitev in izvedbo vedenj, ki na prvi pogled za posameznika niso ekonomična ali smotrna. Zato je smiselno vprašanje, ali imamo morda kakšno nevrološko podlago za takšno vedenje, kot je skrb za potomce.
Ker je starševsko vedenje relativno kompleksno, proučujemo nevrološko povezanost tega vedenja s tradicionalnimi nevrološkimi pristopi, kot so raziskave na temelju lezij (tudi induciranih žariščnih poškodb), z raziskavami na živalih in živalskih modelih, ter s sodobnejšimi tehnikami in pristopi, omenjenimi v nadaljevanju. S tehniko magnetne resonance, imenovano slikanje z difuzijskimi tenzorji (angl. diffusion tensor imaging, DTI), ki izkorišča prosto gibanje vodnih molekul, lahko prikažemo snope in potek živčnih vlaken, kjer je gibanje vodnih molekul zaradi urejenosti vlaken bolj omejeno. To imenujemo anatomska povezljivost. Možganske regije, ki so anatomsko povezane, so praviloma med seboj povezane tudi funkcionalno. Funkcionalno povezljivost regij, in posredno s tem dejavnost nevronov, merimo s spremembo nasičenosti krvi s kisikom (oksigenacijo), s tehniko, imenovano funkcionalno slikanje z magnetno resonanco (angl. functional magnetic resonance imaging, fMRI). Ko spontana aktivnost v dveh regijah pozitivno ali negativno korelira, velja, da so regije med seboj funkcionalno povezane. Vendar pa funkcionalna povezljivost zagotavlja drugačen prikaz kot anatomska povezljivost, saj odraža vpliv obsežnejših večsinaptičnih povezav in povezanih odnosov med regijami možganov. Pri kompleksnih vedenjih, kot je skrb za potomce, moramo zato upoštevati vse naštete vidike proučevanja možganov, preden lahko z gotovostjo trdimo, da smo odkrili, kaj se dogaja v možganih.
Katera možganska področja so ključna za starševsko vedenje?
Poskusi klasičnih lezij pri glodalcih so v to fascinantno vedenje vključili veliko možganskih področij 17, 18. Poenostavljeno shemo predstavim na sliki 1. Eno od področij, ki je v raziskavah dosledno opredeljeno kot bistveno pri skrbi za potomce, je srednje predoptično območje oz. jedro (angl. medial preoptic nucleus, MPOA). Najdemo ga globoko v evolucijsko starem delu možganov, v hipotalamusu 19. Za lažjo predstavo je področje prikazano na sliki 2. Sklepamo, da kljub temu, da raziskave temeljijo na modelu lezij pri glodalcih, ki so inducirane specifične možganske okvare, podobna anatomska področja veljajo pri ljudeh. Področje MPOA je bogato z receptorji za oksitocin, ki je poljudno poznan kot hormon ljubezni 20, z receptorji za steroidne hormone, kot je progesteron, poljudno imenovan tudi hormon nosečnosti 21, 22 in z receptorji za prolaktin in tudi nevroni, ki ga izražajo (prolaktin je poljudno znan kot hormon dojenja) 23. Nič čudnega torej ni, da to območje velja za ključno pri starševskem, predvsem materinskem vedenju.
![Slika 1: Shematski prikaz možganskih področij, ki vplivajo na starševsko vedenje. Prirejeno po Kohl in sod., 2017 [^18].](/esinapsa/media/pics/2019-16/Bregant/slika1.png) Slika 1: Shematski prikaz možganskih področij, ki vplivajo na starševsko vedenje. Prirejeno po Kohl in sod., 2017 18.
Slika 1: Shematski prikaz možganskih področij, ki vplivajo na starševsko vedenje. Prirejeno po Kohl in sod., 2017 18.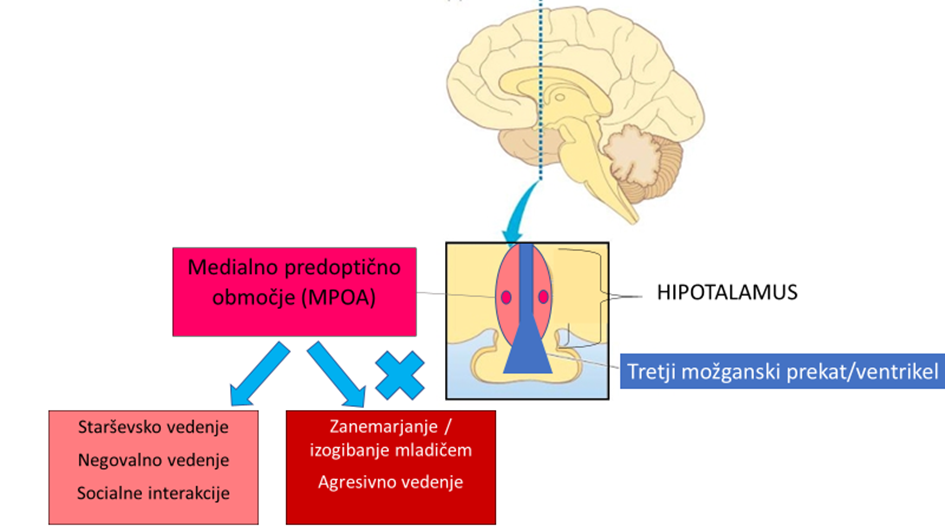 Slika 2: Prikaz ključnega možganskega področja, ki vpliva na starševsko vedenje.
Slika 2: Prikaz ključnega možganskega področja, ki vpliva na starševsko vedenje.Nevroni MPOAGal in njihove povezave
V hipotalamusu, v področju MPOA, so raziskovalci pri miših odkrili posebno skupino nevronov, nevrone MPOAGal 19. Ti nevroni so prisotni pri obeh spolih in so ključni za starševsko obnašanje 17. Teh nevronov je zelo malo, približno 10.000 od skupno 100 milijonov v mišjih možganih, zato domnevamo, da delujejo kot dirigent v orkestru, ki usklajuje različna vedenja oziroma delujejo kot vozlišča, saj je starševstvo tudi pri glodalcih precej kompleksno. Sestavljeno je iz stereotipnih motoričnih rutin, kot je na primer urejanje (lizanje in gladenje) mladičev, njihovo nameščanje v gnezdu in dojenje. Glodalci, ki so starši, se tako vedejo precej dlje časa kot pa drugi odrasli, ki se ta čas ukvarjajo na primer s parjenjem 18.
Bistveno vozlišče za družbeno vedenje je področje MPOA, ki je sestavljeno iz molekularno raznolikih nevronov z razširjenimi projekcijami. V tem območju lahko biokemično identificiramo podskupine nevronov, ki imajo na membranah izražene različne receptorje ter izražajo in v okolico sproščajo različne nevromodulatorje: na primer nevrone MPOAPrl, ki izražajo in imajo tudi receptorje za prolaktin ter so dejavni med nosečnostjo in dojenjem, vplivajo pa tudi na plastičnost možganov, aksioznost in vzburjenje; nevrone MPOANts, ki izražajo nevrotenzin (Nts) in se odzivajo na steroide ter povezujejo z ventralnim tegmentnim območjem (VTA), ter tako tvorijo socialno angažiran krog nagrajevanja 24; ter nevrone MPOAGal , ki izražajo nevropeptid galanin.
Nevroni MPOAGal so organizirani v funkcionalne skupine, od katerih vsaka projicira na drugo področje možganov, pri čemer ima vsaka skupina dostop do vhodnih informacij iz skoraj vseh možganskih področij in je dejavna pri starševskem vedenju 25. Poznamo posamične diskretne skupine teh nevronov, ki so posebej aktivni tekom posamezne faze starševskega vedenja, na primer med porodom. Z uporabo optične fotometrije, ki svetlobno beleži populacijsko aktivnost izbranih nevronov, so se za najbolj dejavne izkazale skupine nevronov, ki so pošiljale nitje v tri področja: periakveduktalno sivino (angl. periaqueductal gray, PAG), sprednje tegmentno območje (angl. ventral tegmental area, VTA) in srednje jedro amigdale (angl. medial amygdaloid nucleus, MeA) 19.
Vpliv spola na starševsko vedenje
Pri sesalcih je za preživetje potomcev ključna starševska, predvsem materina, skrb. Ključno vozlišče je MPOA, na katerega posredno vplivajo hormonska nihanja in vnos zaznavnih informacij. Naraščajoče ravni estradiola, oksitocina, prolaktina in placentnega laktogena skupaj z nenadnim zmanjšanjem progesterona, sprožijo aktivacijo tipičnega materinega vedenja takoj po rojstvu oziroma kotitvi 26. MPOA pri tem tvori kritično vez, saj združuje vrsto hormonskih in senzoričnih vnosov v tako imenovano »materinsko omrežje« 24.
V materinskem omrežju lahko razločimo specifične skupine nevronov, predvsem izstopajo na prolaktin odzivni nevroni, katerih projekcijsko nitje se razprostira po vseh možganih 25. Po načrtnem izbijanju receptorjev prolaktina na teh nevronih odraslih samic, so miši lahko zanosile in normalno skotile, a so kmalu opustile legla in niso izkazovale normalnega materinskega obnašanja. Lahko sklepamo, da je za vzpostavitev in vzdrževanje starševskega vedenja delovanje prolaktina v področju MPOA ključno.
Običajno samice ne izražajo telesne agresije v tolikšni meri kot samci. Izjema je materinska zaščita potomcev, ki pa jo pojasnjujemo z delovanjem prolaktina in ne z delovanjem testosterona. Testosteron ima sicer pozitivne korelacije z agresivnostjo in negativne korelacije s kontekstom starševstva 21, 27. Praviloma imajo očetje nižjo izhodiščno vrednost testosterona kot moški ne-očetje 27. Le malo raziskav pa je primerjalo izhodiščne ravni testosterona med materami in ženskamii, ki to niso. Nekaj študij kaže, da velja podobno in imajo matere nižje vrednosti testosterona kot samice, ki niso matere. To je sicer kontradiktorno z agresivnim vedenjem samic-mater, ki pa ga lahko pojasnimo s tem, da je vezano na delovanje prolaktina in ne na običajno delovanje testosterona. Razlike med spoloma bi lahko pojasnili s tem, da je reproduktivni uspeh samcev pogosto omejen z dostopom do samic, medtem ko je reproduktivni uspeh samic omejen z dostopom do virov preživetja, kot je hrana. Samica v nosečnosti in ob dojenju investira tudi metabolno, zato je njena agresija v tem obdobju evolucijsko razumljiva.
Pri miših so spolno neizkušeni samčki običajno agresivni do mladičev. Pri njih opažamo infanticidno vedenje do mladičev, ki niso njihovi. Starševsko vedenje se pri njih izrazi šele nekaj tednov po parjenju 28. Aktivacija nevronov MPOAGal, ki projicirajo v področje PAG, pri njih zavre siceršnjo agresijo do mladičev. Ob aktivaciji teh nevronov pride tudi do povečanega negovalnega vedenja do mladičev pri obeh spolih 28. Nasprotno pa velja za aktiviranje nevronskega bazena, ki projicira v področje VTA, ki sicer velja za pomembno področje pri motiviranju in nagrajevanju 29, 30. Ker so starši posebej motivirani za interakcijo z mladiči, so raziskovalci vstavili prehodno pregrado med preskusno živaljo in mladiči. V tem preprostem testu z omejevanjem je aktivacija skupine nevronov, ki projicira v področje VTA, znatno povečala pogostost prehajanja odraslih živali v predelek z mladiči, kar kaže, da ta signalizacija nadzoruje motivacijo za interakcijo z mladiči. Zanimivo pa je bilo, da aktiviranje nevronov MPOAGal, ki projicirajo v področje amigdale, ni vplivalo niti na interakcijo med mladiči niti na interakcijo staršev z njimi, je pa to zavrlo interakcije odraslih miši med sabo, neodvisno od spola 29. Ta skupina nevronov bi torej lahko imela posredno vlogo pri spodbujanju starševstva z zatajevanjem socialnega vedenja, ki bi lahko vodilo v parjenje.
Agresija napram mladičem je poznana pri več živalskih vrstah. Ko samec postane nov spolni partner matere, se verjetnost za agresijo proti njenim mladičem znatno poveča. Gre za neke vrste evolucijski boj, saj s tem novi partner poveča lasten genski bazen, saj matere ubitega mladiča prenehajo dojiti, vzpostavi se ovulacija in mati se lahko ponovno razmnožuje 31. Pri infanticidnem vedenju je bolj vpleteno delovanje nevronov iz amigdale in stresno okolje, vendar pa je mehanizem zaenkrat slabo pojasnjen. Takšno vedenje poznamo tudi pri ljudeh. Pri ljudeh je sicer družinska struktura, velja za družine v ZDA, kjer je bila raziskava narejena, najpomembnejši dejavnik tveganja pri zlorabi otrok in detomoru 32. Otroci, ki živijo s svojimi naravnimi (biološkimi) starši, so izpostavljeni nizkemu tveganju za zlorabo. Tveganje se poveča, če otroci živijo s samo enim od bioloških staršev. Otroci, ki živijo brez enega od staršev, imajo 10-krat večjo verjetnost zlorabe kot otroci, ki živijo z obema biološkima staršema. Otroci, ki živijo z biološkim staršem, ki ima novega partnerja, pa so v največji nevarnosti: 20-krat bolj je verjetno, da so žrtve zlorabe otrok kot otroci, ki živijo z obema biološkima staršema 32.
Zaključek
Iz raziskav sklepamo, da v področju hipotalamusa obstajajo posebne skupine nevronov MPOAGal, ki glede na projekcijsko mesto, določajo specifične vidike starševstva. Ker pa je ta skupina nevronov majhna, je pravo vprašanje, kako dosežejo tako močne vplive na kompleksno vedenje, kot je starševstvo. Pri ljudeh na starševsko skrb vplivajo tudi stres, duševne bolezni in druga družbeno pogojena stanja, ki so v novejšem času v porastu. Poporodna depresija in/ali tesnoba na primer prizadeneta skoraj petino vseh novih mater 33. Z raziskovanjem starševskega vedenja drugih živali in s pomočjo nevroznanosti lahko pridobimo nekaj uvida v lastno delovanje in tako poskušamo razumeti izjemno spremenljivo odzivnost človeških staršev.
-
___
-
Clutton‐Brock TH. The Evolution of Parental Care. 1991. Princeton University Press, Princeton, New Jersey, USA. ↩
-
Gross MR. The evolution of parental care. The Quarterly Review in Biology. 2005;80:37–45. ↩
-
Carlisle TR. Brood success in variable environments: implications for parental care allocation. Animal Behaviour. 1982;30:824–836. ↩
-
Davis JN, Todd PM, Bullock S. Environment quality predicts parental provisioning decisions. Proceedings Of The Royal Soceity Of London. Series B: biological Sciences. 1999;266:1791–1797. ↩
-
Reynolds JD, Goodwin NB, Freckleton RP. Evolutionary transitions in parental care and live bearing in vertebrates. Philosophical Transactions of the Royal Societ of London B. 2002;357:269–281. ↩
-
Lynn SE. Endocrine and neuroendocrine regulation of fathering behavior in birds. Hormones and Behavior. 2016;77:237–248. ↩
-
Summers K, Weigt LA, Boag P, Bermingham E. The evolution of female parental care in poison frogs of the genus Dendrobates: evidence from mitochondrial DNA sequences. Herpetologica. 1999;55:254–270. ↩
-
Hrdy SB. Variable postpartum responsiveness among humans and other primates with “cooperative breeding”: A comparative and evolutionary perspective. Hormones and Behavior. 2016;77:272–283. ↩
-
Young MC. The Guinness Book of World Records. 1998. Bantam Books. ↩
-
Clay MM. Feodor Vassilyev: a case of remarkable fecundity. Quadruplets and Higher Multiple Births. 1989. London: Mac Keith Press. ↩
-
Zerjal T, Xue Y, Bertorelle G, Wells RS, Bao W, et al. The genetic legacy of the Mongols. Am J Hum Genet. 2003;72(3):717–721. ↩
-
Oberzaucher E, Grammer K. The case of Moulay Ismael–fact or fancy? PLoSONE9. 2014; e85292. ↩
-
Sear R, Lawson DW, Kaplan H, Shenk MK. Understanding variation in human fertility: What can we learn from evolutionary demography? Philosophical Transactions of The Royal Society B Biological Sciences. 2016;371(1692): 20150144. ↩
-
Kaplan H. Evolutionary and Wealth Flows Theories of Fertility: Empirical Tests and New Models. Population and Development Review. 1994;20(4):753-791. ↩
-
Capital group 2018. American funds. 2018. How Much Does It Cost to Raise a Child. Dostopno na: https://www.americanfunds.com/individual/planning/college-savings/cost-of-raising-a-child.html (7.1.2019). ↩
-
Bratanič J, Košir K. 100.000 evrov od zibke do srednje šole. Za enega otroka! Delo, Svet kapitala. 28. september 2018. Dostopno na: https://svetkapitala.delo.si/trendi/100-000-evrov-od-zibke-do-srednje-sole-za-enega-otroka-6583 (7.1.2019). ↩
-
Dulac C, O’Connell LZ, Wu Z. Neural control of maternal and paternal behaviors. Science. 2014;345(6198):765-770. ↩
-
Kohl J, Autry AE, Dulac C. The neurobiology of parenting: A neural circuit perspective. Bioessays. 2017;39(1):1-11. ↩
-
Kohl J. Circuits for care: A small population of hypothalamic neurons orchestrates parenting behaviors. Science. 2018;362(6411):168-169. ↩
-
Nishimori K, Sato K, Hidema S, Yoshida M, Mizukami H. Oxytocin Receptor-Expressing Neurons and Nuclei in the Regulation of Social Behaviors. Interdisciplinary Information Sciences. 2015;21(3):283–288. ↩
-
Oberlander JG, Henderson LP. The Sturm und Drang of anabolic steroid use: angst, anxiety, and aggression. Trends Neurosci. 2012;35(6):382-392. ↩
-
Barbosa-Vargas E, Pfaus JG, Woodside B. Sexual behavior in lactating rats: role of estrogen-induced progesterone receptors. Horm Behav. 2009;56(2):246-253. ↩
-
Consiglio AR, Bridges RS. Circulating prolactin, MPOA prolactin receptor expression and maternal aggression in lactating rats. Behav Brain Res. 2008;197(1):97-102. ↩
-
Numan M, Woodside B. Maternity: Neural mechanisms, motivational processes, and physiological adaptations. Behav Neurosci. 2010;124:715–741. ↩
-
Brown RSE, Aoki M, Ladyman SR, Phillipps HR, Wyatt A, Boehm U, et al. Prolactin action in the medial preoptic area is necessary for postpartum maternal nursing behavior. PNAS. 2017;114(40):10779-10784. ↩
-
Bridges RS. Neuroendocrine regulation of maternal behavior. Front Neuroendocrinol. 2015;36:178–196. ↩
-
Gray PB, Anderson KG. Fatherhood. Evolution and Human Paternal Behavior. 2012. Harvard University Press, USA. ↩
-
vom Saal FS. Time-contingent change in infanticide and parental behavior induced by ejaculation in male mice. Physiol Behav. 1985;34(1):7-15. ↩
-
Salamone JD, Correa M. The mysterious motivational functions of mesolimbic dopamine. Neuron 2012;76(3):470-485. ↩
-
McHenry JA, Otis JM, Rossi MA, Robinson JE, Kosyk O, Miller NW. Hormonal gain control of a medial preoptic area social reward circuit. Nat Neurosci. 2017;20(3):449-458. ↩
-
Hausfater G. Infanticide: Comparative and Evolutionary Perspectives. Current Anthropology. 1984;25(4):500–502. ↩
-
Sedlak AJ, Mettenburg J, Basena M, Petta I, McPherson K, Greene A, et al. Fourth National Incidence Study of Child Abuse and Neglect (NIS–4): Report to Congress, Executive Summary. Washington, DC: U.S. Department of Health and Human Services, Administration for Children and Families, 2010, USA. ↩
-
Wisner KL, Sit DKY, McShea MC, Rizzo DM, Zoretich RA, Hughes CL, et al. Onset Timing, Thoughts of Self-harm, and Diagnoses in Postpartum Women With Screen-Positive Depression Findings. JAMA Psychiatry. 2013;70(5):490–498. ↩
doc. dr. Tina Bregant, dr. med., spec. pediatrije, spec. FRM
Center za izobraževanje, rehabilitacijo in usposabljanje – CIRIUS Kamnik
Prejeto: 15.1.2019
Objavljeno: 25.3.2019